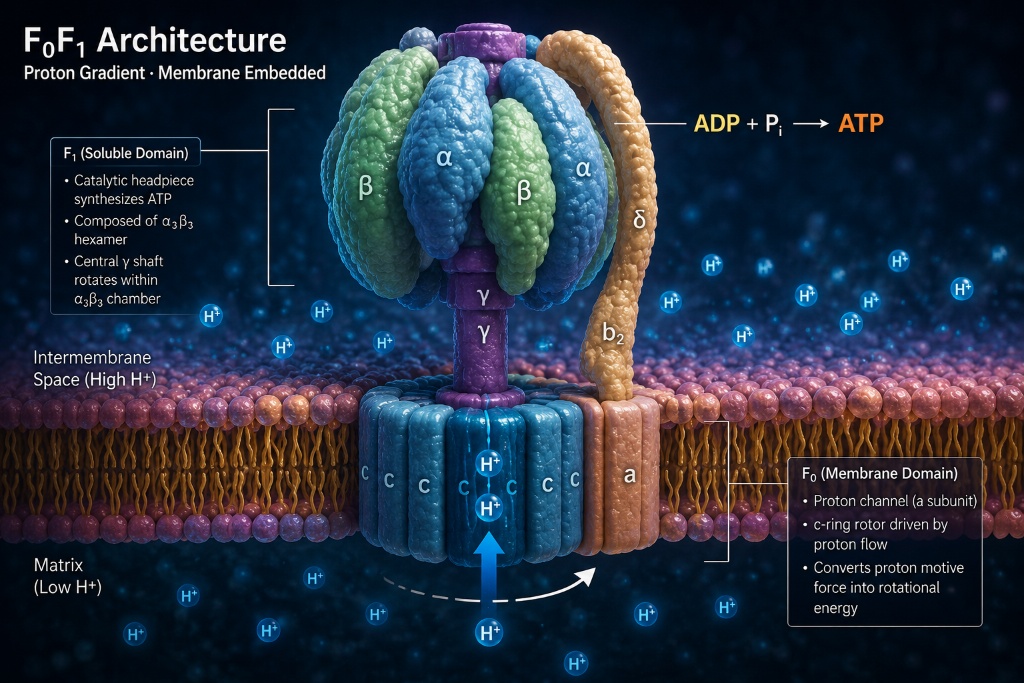
 F₀F₁ Architecture · Proton Gradient · Membrane Embedded
F₀F₁ Architecture · Proton Gradient · Membrane Embedded
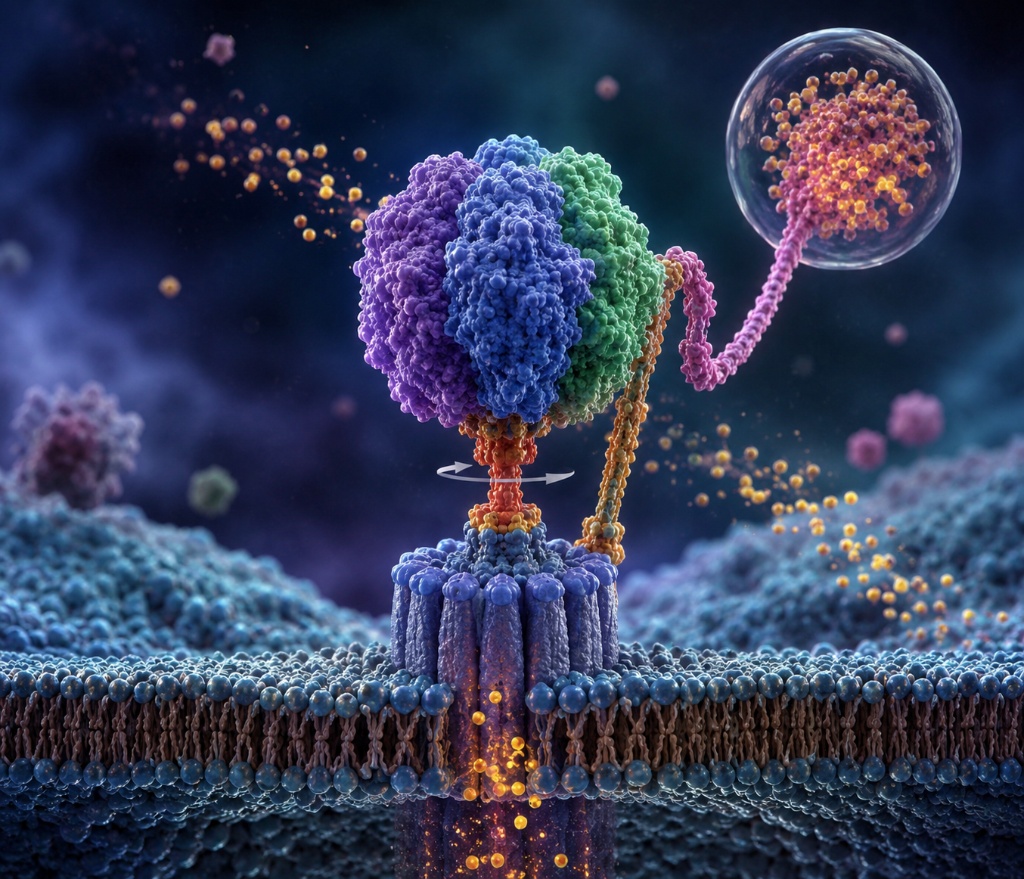 ATP Synthase · Rotary Nanomotor · >95% Efficiency
ATP Synthase · Rotary Nanomotor · >95% Efficiency
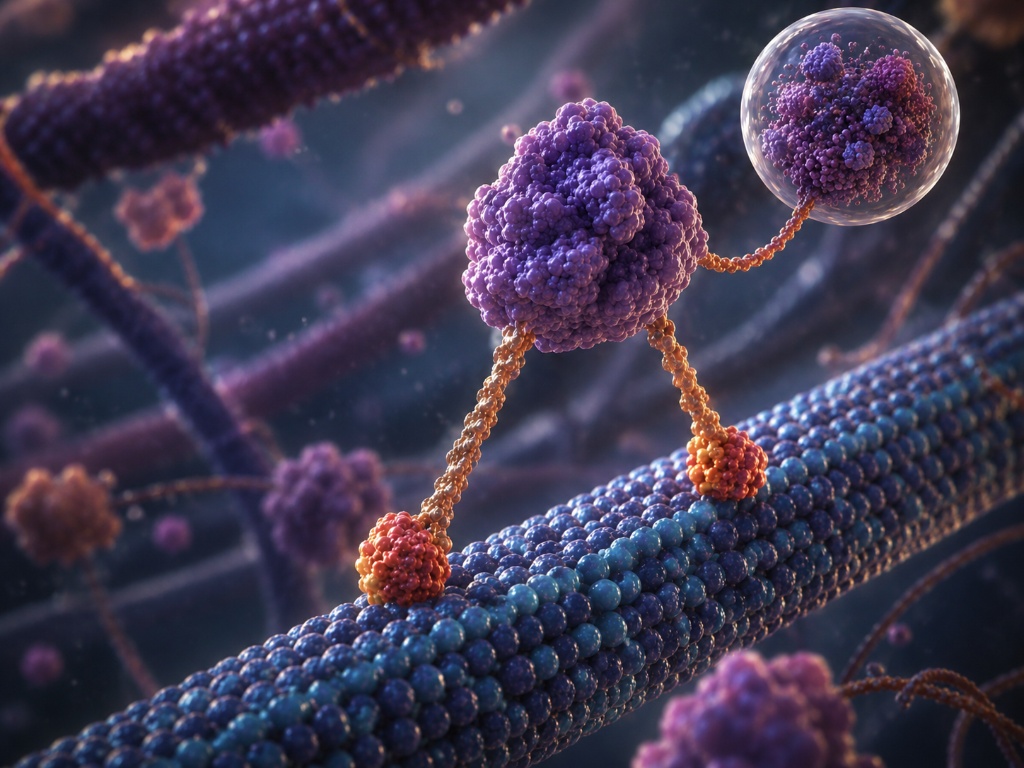 Kinesin · Molecular Walker · 8nm Steps
Kinesin · Molecular Walker · 8nm Steps
Every living cell runs twelve simultaneous, interdependent communication systems. None is redundant. Remove one and the cell dies. All twelve were present in the first cell.
This is not evolution. This is engineering.The argument from biological coherence is not about any single system. It is about the simultaneous requirement of all twelve. Each system assumes the others are already functional. DNA assumes the ribosome exists to read it. The ribosome assumes the RNA processing machinery is already in place. Error correction assumes the replication machinery exists to make errors. Epigenetic regulation assumes the genetic code exists to regulate.
The undirected-process narrative requires that complex systems arise in stages, with each intermediate stage conferring a survival advantage. There are no intermediate stages for systems that only work when complete. You cannot have half a genetic code. You cannot have partial error correction. You cannot have a cell with six of the twelve systems and wait for a blind search process to supply the other six. Without all twelve, there is no cell. What biological systems do exhibit — adaptation to external conditions, optimization within a range — is design responding to environment, not design arising from it.
The first cell that functioned already had all twelve. The question is not whether they arose by chance. The question is whether chance is even a coherent explanation for a system of this specification density.
F₀F₁ Architecture · Proton Gradient · Membrane Embedded
ATP Synthase · Rotary Nanomotor · >95% Efficiency
Kinesin · Molecular Walker · 8nm Steps
Each one alone is remarkable. Together they are conclusive.
The four-letter digital code in every cell specifies protein assembly with precision that surpasses every human-engineered data system. The codon-to-amino-acid mapping is arbitrary — it could map differently, but it doesn't. Arbitrariness is the signature of language. Language requires a speaker.
Deep Dive → SYSTEM · 02 Epigenetic Code A second code written on top of the first. Context-dependent. Heritable. Reversible.Chemical marks on the DNA strand control which genes are read without changing the sequence. This is a second language operating on top of the genetic language — with its own alphabet, grammar, and inheritance rules. Two complete information systems, layered.
Deep Dive → SYSTEM · 03 Histone Code A third code — chemical modifications on the protein spools that package DNA.Histones are not passive packaging. They carry a combinatorial chemical code — acetylation, methylation, phosphorylation — that regulates gene expression. Three complete, interdependent information systems before the ribosome reads a single codon.
Deep Dive → SYSTEM · 04 DNA Methylation Cytosine marks that silence genes across cell generations. Tissue identity in chemical form.Methylation patterns determine cell identity — why a liver cell and a neuron carry identical DNA but behave entirely differently. The pattern is heritable across cell division. It is tissue memory encoded in chemistry.
Deep Dive → SYSTEM · 05 Glycocalyx — Sugar Code A fourth information layer — sugar chains on the cell surface encoding identity and communication.The glycocalyx is a forest of complex sugar chains coating every cell. It encodes cell identity, mediates immune recognition, guides development, and enables cell-to-cell communication. A fourth complete information system, expressed at the cell surface.
Deep Dive → SYSTEM · 06 Enzymatic Expression Protein machines that catalyse reactions 10⁶ to 10¹² times faster than uncatalysed rates.Enzymes are specified molecular machines — each exquisitely matched to its substrate, each catalysing a precise reaction. What appears as environmental optimization is a pre-specified design response, not undirected selection producing novelty. Without enzymes, life's chemistry runs too slowly to sustain life. The first cell required thousands of working enzymes simultaneously. Axe's estimate: 1 in 10¹⁵⁰ random sequences is functional.
Deep Dive → SYSTEM · 07 RNA Regulation microRNA, lncRNA, siRNA — a vast layer of gene expression control once called "junk."The ENCODE project found that 80% of the genome formerly dismissed as junk has biochemical function. Thousands of RNA molecules regulate gene expression at the post-transcriptional level. The genome is not a parts list. It is an orchestrated score.
Deep Dive → SYSTEM · 08 Ribozymes — RNA Machines RNA molecules that act as enzymes. The ribosome itself is a ribozyme.The ribosome — the machine that translates DNA code into proteins — is itself a ribozyme. It uses RNA to catalyse peptide bond formation. RNA both carries information and performs catalysis. The same molecule. This is not a simple system.
Deep Dive → SYSTEM · 09 Error Correction 1 error per 10⁹ base pairs. Proofreading, mismatch repair, nucleotide excision — all simultaneous.DNA replication has an error rate lower than any human-engineered information system. Three independent proofreading mechanisms operate simultaneously. Without them, the genome degrades in a single generation. Error correction presupposes the replication machinery it corrects.
Deep Dive → SYSTEM · 10 Cell Differentiation One genome. 200+ cell types. The same code read differently in every tissue.A liver cell and a neuron carry identical DNA. Differentiation is the process by which the same information produces radically different machines. It requires the epigenetic, histone, and methylation systems all functioning correctly before the first division. Identity is specified before function begins.
Deep Dive → SYSTEM · 11 Protein Folding Levinthal's Paradox: a protein cannot find its correct fold by random search in the age of the universe.A 150-residue protein has 10³⁰⁰ possible conformations. Random sampling at 10¹³ per second would take longer than the age of the universe to find the correct fold. Yet proteins fold in milliseconds. Chaperone proteins guide folding — but they too must be correctly folded first. The problem is recursive.
Deep Dive → SYSTEM · 12 Cell Signaling 12 simultaneous signaling networks — hormonal, electrical, chemical, mechanical, epigenetic.A single cell simultaneously operates 12 distinct signaling networks. Each assumes the others are functioning. No evolutionary story explains how 12 interdependent systems arose independently and then integrated. The first cell that worked already had all twelve.
Deep Dive →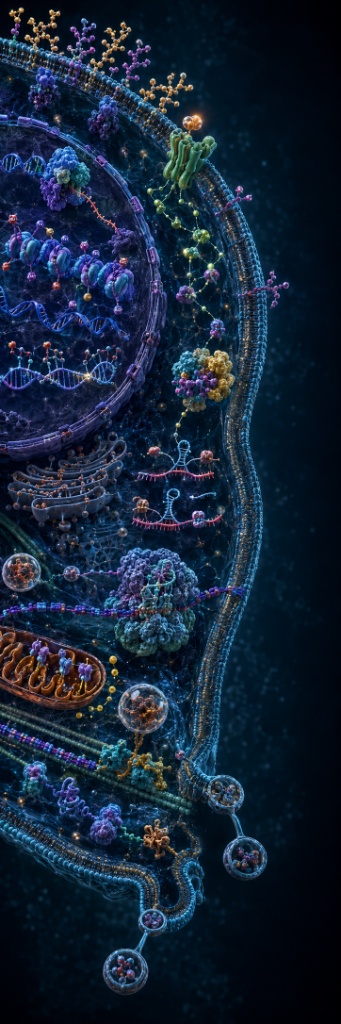
Select any system to expand the argument. The image to the left shows every one of these running simultaneously right now.
3.2 billion letters. Error rate: 1 per billion. Arbitrary to chemistry.
A second code written on top of the first. Context-dependent. Heritable. Reversible.
A third code — chemical modifications on the protein spools that package DNA.
Cytosine marks that silence genes across cell generations. Tissue identity in chemical form.
A fourth information layer — sugar chains on the cell surface encoding identity.
Protein machines that catalyse reactions 10⁶ to 10¹² times faster than uncatalysed rates.
microRNA, lncRNA, siRNA — a vast layer of gene expression control once called "junk."
RNA molecules that act as enzymes. The ribosome itself is a ribozyme.
1 error per 10⁹ base pairs. Three independent proofreading mechanisms — all simultaneous.
One genome. 200+ cell types. The same code read differently in every tissue.
A 150-residue protein has 10³⁰⁰ possible folds. Random search takes longer than the universe.
12 simultaneous signaling networks — hormonal, electrical, chemical, mechanical, epigenetic.
"An honest man, armed with all the knowledge available to us now, could only state that in some sense, the origin of life appears at the moment to be almost a miracle, so many are the conditions which would have had to have been satisfied to get it going."
— Francis Crick · co-discoverer of DNA · Life Itself, 1981The 12 systems are one argument. There are two others — the motor, and the universe. Each one alone is conclusive. Together they are inescapable.