Molecular
Machines That
Run Every Cell
Enzymes don't merely help reactions happen — they make impossible chemistry possible, with tolerances no human engineer would claim. The masterclass: ATP synthase, a literal rotary nanomotor spinning at 120 RPM, synthesizing the energy currency of all life from a proton gradient. In 1997, Masasuke Yoshida's team directly filmed it rotating. The viable window: ±0.3 pH units. Step outside it — the motor stops. The cell dies.
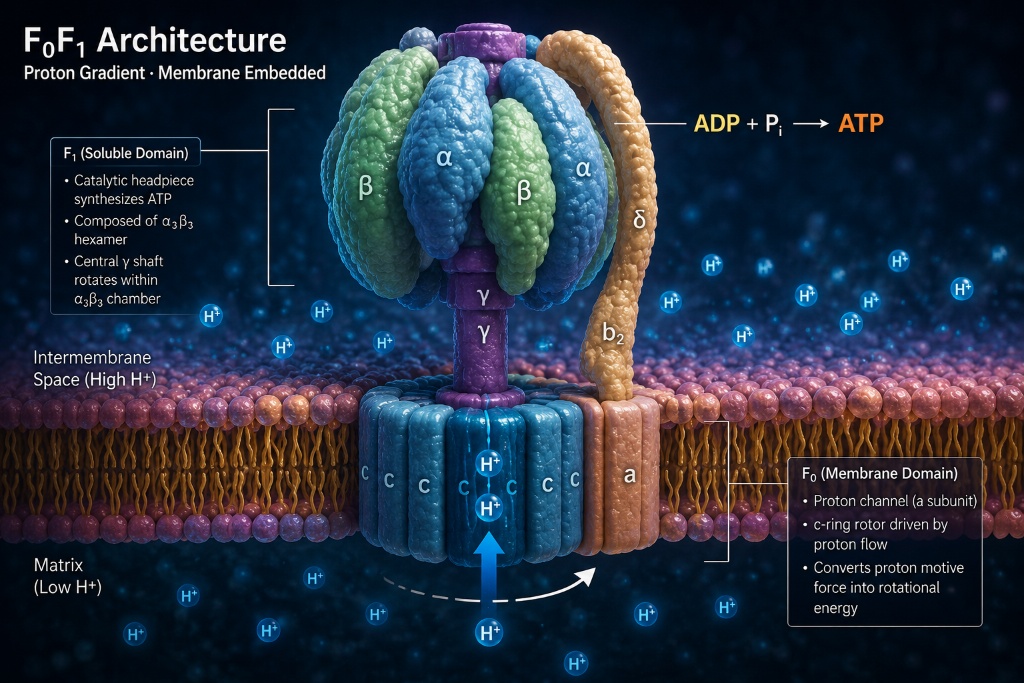 F₀F₁ Architecture · Proton Gradient · Membrane Embedded
F₀F₁ Architecture · Proton Gradient · Membrane Embedded
Not catalysts. Molecular machines.
Every chemical reaction in a living cell is carried out by an enzyme. Not some reactions — every one. DNA is copied by a polymerase enzyme. RNA is built by an enzyme. Proteins are assembled by a ribosome — itself a complex of enzymes. Waste is cleared by enzymes. Signals are sent and received by enzymes. The entire metabolic map of a cell is a network of enzyme-catalyzed transformations running in sequence, in parallel, in feedback loops, at the exact rates life requires. Remove any single enzyme class and the cascade that depended on it fails.
What makes this remarkable is not the number of enzymes — it is their precision. An enzyme active site fits its substrate the way a key fits a lock — except that the key bends the lock slightly as it enters, inducing a conformational change that triggers the reaction. Induced fit. Wrong substrate: no fit, no change, no reaction. Right substrate: catalysis proceeds with rate enhancements of up to 1017-fold over the uncatalyzed reaction. That number is not a rounding error. That is the difference between chemistry that happens in milliseconds and chemistry that would take longer than the age of the universe.
II · The MasterclassNot a metaphor. A machine.
ATP synthase is a molecular motor embedded in the inner mitochondrial membrane. It consists of two rotary units — F₁ (above the membrane, in the mitochondrial matrix) and F₀ (embedded in the membrane) — connected by a central rotating shaft. As protons flow through F₀ from the intermembrane space into the matrix, the pressure drives the shaft to rotate. Each full rotation of the shaft causes F₁ to synthesize three molecules of ATP — adenosine triphosphate, the energy currency of all life.
The 1997 confirmation by Noji, Yasuda, Yoshida and Kinosita used a fluorescent actin filament attached to the F₁ rotor. Under an optical microscope, the filament rotated in discrete 120° steps — one step per ATP hydrolyzed. The experiment left no interpretive room: this is a mechanical rotary device, operating by physical torque, at the molecular scale. No prior model of biology had prepared the scientific community for this.
Rotations per minute under physiological proton gradient (ΔpH 1.4 across membrane)
ATP molecules synthesized per full 360° rotation — one per 120° step
ATP molecules synthesized in your body every second — 40 kg of ATP per day in a 70 kg adult
Eight distinct subunits. One coordinated machine.
Embedded in the inner mitochondrial membrane. A ring of 8–15 c-subunits (species-dependent) that rotates as protons pass through the interface with the a-subunit. The stoichiometry of the c-ring determines the thermodynamic efficiency of ATP synthesis.
A hexamer of alternating α and β subunits (α₃β₃). The three β subunits are the catalytic sites. As the central γ shaft rotates, each β subunit cycles through binding (ADP+Pi), catalysis (ATP formation), and release states — the binding-change mechanism proposed by Paul Boyer (Nobel 1997).
The rotating axle connecting F₀ to F₁. Its asymmetric shape is what converts c-ring rotation into the conformational changes in β subunits that drive ATP synthesis. The shaft rotates inside the α₃β₃ hexamer like a cam in a bearing.
The b₂δ peripheral stalk holds F₁ stationary relative to the membrane while F₀ and the γ shaft rotate. Without this structural constraint, rotation would spin the entire F₁ head rather than driving conformational change. The stator is a mechanical necessity with no simpler functional precursor.
Drag the sliders. Watch the motor stop.
ATP synthase operates within extraordinarily narrow parameter windows. The proton motive force, the membrane potential, the ADP/ATP ratio, and the rotational torque all must fall within tight bounds for the motor to function. This is not a robust system with generous margins. It is a precision instrument operating at the edge of thermodynamic viability. Adjust any parameter below and observe the consequence.
At ΔpH 1.4: proton motive force drives steady 120 RPM rotation. ATP synthesis at full capacity.
At −140 mV: combined with ΔpH, total proton motive force ≈ −200 mV. Sufficient for ATP synthesis.
At 80% ADP: active site occupation is high. Motor operates near maximal throughput.
At 37°C: optimal enzymatic rate. Thermal energy sufficient for proton translocation without denaturation.
An unguided process cannot produce a precision rotary motor.
The Goldilocks explorer above demonstrates the problem in physical terms. ATP synthase does not merely require the right components. It requires the right components assembled in the right configuration, operating in the right electrochemical environment, within the right temperature range, with the right substrate availability — simultaneously, from the first moment of function. Remove the stator and the catalytic head spins as a unit — no ATP. Remove the c-ring and there is no rotor — no ATP. Remove the proton gradient and the thermodynamic driving force is absent — no ATP. There is no functional reduced version.
This is what Michael Behe called irreducible complexity — not as a philosophical claim but as a straightforward engineering observation. A system is irreducibly complex if the removal of any single component causes complete loss of function. ATP synthase meets this criterion at the level of individual subunits. It meets it at the level of its electrochemical environment. The system does not merely require assembly. It requires simultaneous assembly of all of its parts in the correct geometric relationship, in an environment that also had to be pre-established for the motor to function at all.
The evolutionary narrative requires that a functional ATP synthase assembly arose from non-functional precursors through a continuous series of selectable steps. This requires that each intermediate be selectable — i.e., provide a fitness advantage. But an incomplete ATP synthase synthesizes no ATP. A c-ring without the F₁ head is not "a little bit functional." It is non-functional. The selective pressure for each component appears only when all components are present. This is the problem that no incremental model has resolved.
Read at Nature ↗ · Google Scholar ↗ · Behe (1996) — Darwin's Black Box ↗