The Information Crisis
Every time a human cell divides, it must copy 3.2 billion base pairs of DNA with extraordinary accuracy. A single uncorrected error in the wrong gene causes cancer. Thousands of errors cause cell death. The raw error rate of DNA polymerase — the molecule that does the copying — is approximately 1 mistake per 100,000 base pairs.
That rate, if uncorrected, would produce roughly 64,000 errors per cell division. A human being has approximately 37 trillion cells. The math is unambiguous: uncorrected replication is incompatible with biological complexity.
In 1971, German physicist and chemist Manfred Eigen (b. 1927) published the mathematical framework that made this problem precise. He proved that there exists a sharp threshold — the error threshold — above which genetic information cannot be stably maintained. Cross the threshold from below, and the genome dissolves into random noise: what he called error catastrophe.
"There is a maximum genome length compatible with stable inheritance, determined by the product of genome length and error rate per base. Above this threshold, the master sequence is lost."
Three Correction Systems
The cell operates three independent, sequential error-correction systems. Each addresses a different class of mistake at a different stage of the replication process. All three are required; removing any one causes a measurable increase in mutation rate and, in humans, cancer predisposition syndromes.
DNA Pol III has a built-in 3'→5' exonuclease domain. After inserting each base, it pauses to verify geometry. Incorrect bases stall the active site — the exonuclease then excises and re-inserts. Reduces error rate by 100-fold, from 10⁻⁵ to 10⁻⁷.
After replication, MutS scans for geometric distortions in the double helix — bulges from mispaired bases. MutL recruits MutH, which cleaves the newly synthesized strand (identified by delayed GATC methylation). The error is excised and re-synthesized. Reduces rate by 1,000-fold.
DNA glycosylases patrol the genome for specific chemical damage: oxidized guanine (8-oxoG), deaminated cytosine (uracil), and alkylated bases. Each glycosylase recognizes one type of lesion, flips the base out of the double helix, and cleaves it — triggering gap-fill synthesis. Continuous process, not replication-coupled.
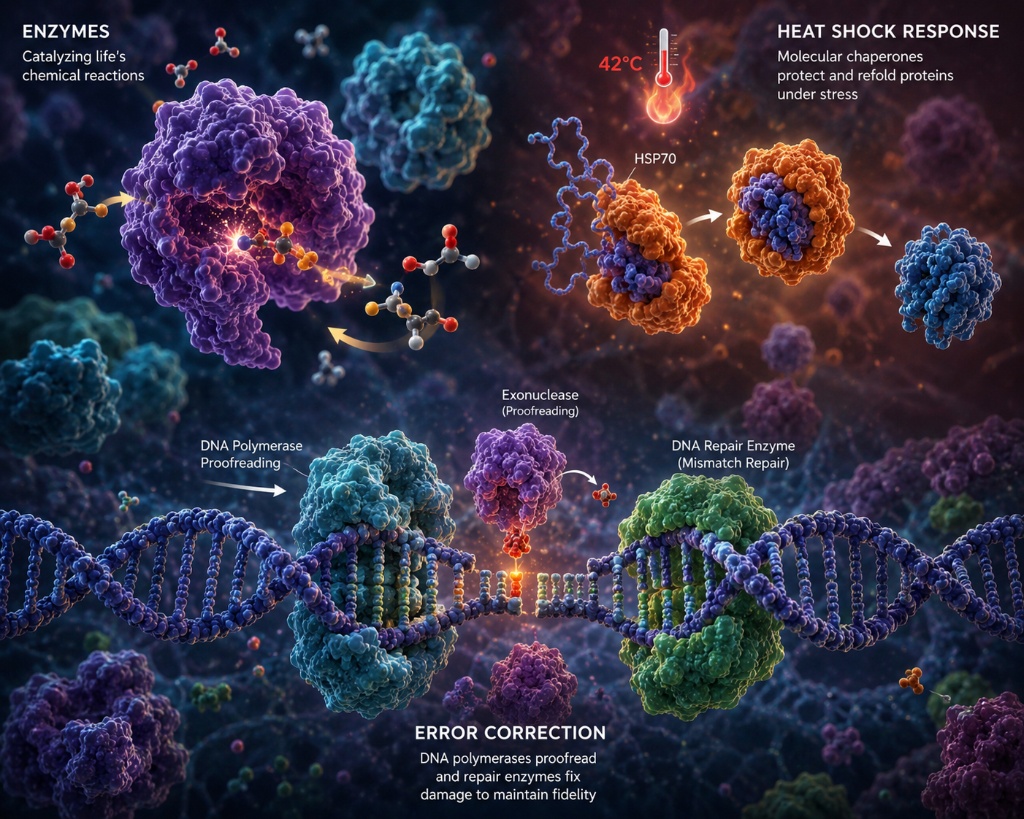
Together, these three systems achieve a combined error reduction of approximately 10⁷-fold — reducing the raw polymerase error rate of 10⁻⁵ to a final net rate of approximately 10⁻⁹ to 10⁻¹⁰ per base pair per replication. For a 3.2-billion base pair genome, this translates to fewer than 1–3 new mutations per cell division on average.
The astonishing detail: these three systems are mechanistically independent. They use different proteins, different recognition mechanisms, different repair pathways. They are not a redundant system where one could evolve in the absence of others. The absence of any one causes hereditary cancer syndromes: Lynch syndrome (MMR deficiency), xeroderma pigmentosum (NER deficiency), MUTYH-associated polyposis (BER deficiency).
III. The Goldilocks ExplorerThe Error Threshold Window
Eigen's theorem specifies a precise mathematical relationship between mutation rate, genome length, and information maintenance. Move any slider below and explore how narrow the viable zone is. The parameters are taken from Eigen's 1971 paper and subsequent empirical measurement.
Survival Probability
What This Requires
The three correction systems did not appear sequentially in evolutionary history — each requires the others to function at all. Without proofreading, the raw error rate is too high for MMR to handle. Without MMR, the residual error rate after proofreading is too high for a large genome. Without BER, chemical damage accumulates between replications faster than either system can address.
Eigen's math is merciless: the error threshold for a genome of human complexity (~3.2 × 10⁹ base pairs) requires a net error rate no worse than approximately 10⁻⁹. With only polymerase proofreading, the best achievable rate is ~10⁻⁷ — two orders of magnitude above what the genome requires. All three systems, working together, achieve the necessary 10⁻⁹ to 10⁻¹⁰.
This is not a case of redundant backup systems. It is a case of irreducible minimum specification: the information storage capacity of the genome is physically impossible without all three layers of correction operating simultaneously. The question of how the information arose when the storage system required to maintain it was not yet in place is precisely Eigen's Paradox — and it remains unresolved.