The Fold Problem
Every protein is a linear sequence of amino acids — typically 100 to 1,000 residues long. When that chain is released from the ribosome, it must achieve a specific three-dimensional fold to function. The correct fold is determined entirely by the amino acid sequence. The same sequence always produces the same fold, across any cell, any organism, any temperature within the viable range.
In 1969, Cyrus Levinthal (b. 1922) calculated the consequences of random search for the native fold. A typical protein has approximately 10 rotatable bonds, each with 3 stable positions — yielding 3¹⁰⁰ ≈ 5 × 10⁴⁷ possible conformations. If the protein sampled each conformation for just 10⁻¹³ seconds (the fastest possible bond rotation), finding the correct fold by random search would take 10²⁷ years — roughly 10¹⁷ times the age of the universe.
Proteins fold in milliseconds. The random search time is 10²⁷ years. Something guides the fold — and that guidance is encoded in the amino acid sequence itself, in ways we are still learning to read.
The actual fold time is microseconds to seconds. The explanation — the energy landscape funnel — was developed by Onuchic, Wolynes, and colleagues in the 1990s. The native state is at the bottom of a funnel in energy space; the protein doesn't search randomly, it rolls downhill. But the funnel itself is a highly specific property of the amino acid sequence — only sequences that produce a smooth, funnel-shaped energy landscape fold reliably.
Possible conformations for a 100-residue protein (3¹⁰⁰). Random search time: 10²⁷ years. Actual fold time: milliseconds. The gap between these numbers is Levinthal's Paradox.
Small proteins (50–100 aa): microseconds. Large domains: milliseconds. The energy landscape funnel guides the search — but only for sequences that produce smooth funnels.
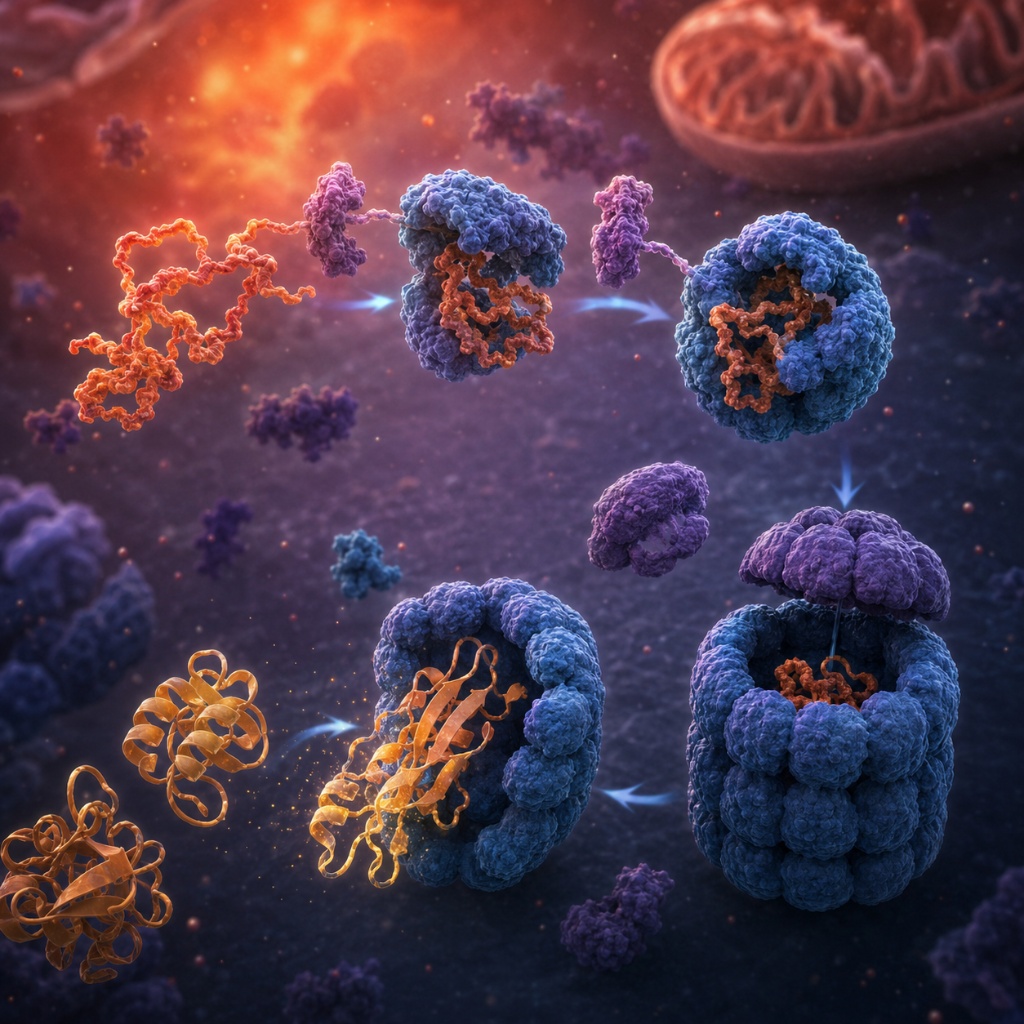
30% of proteins require molecular chaperones (Hsp70, GroEL/GroES) to reach native state in vivo. Chaperones prevent premature aggregation, provide protected folding cavities, and use ATP hydrolysis to actively unfold misfolded intermediates.
The Folding Environment Window
Protein folding is exquisitely sensitive to its physical-chemical environment. The energy landscape funnel that guides folding is distorted by temperature, pH, osmolarity, and crowding. Outside narrow windows on each parameter, the funnel flattens, misfolding occurs, and aggregates form.
Yield
The Information in the Sequence
Levinthal's Paradox is not solved by the energy landscape — it is restated at a deeper level. The energy funnel guides folding efficiently, but the funnel exists only for amino acid sequences that encode it. Most random amino acid sequences do not produce smooth folding funnels. They produce flat or rugged landscapes, where folding is slow, unreliable, or leads to aggregates.
The question shifts from "how does the protein find the fold?" to "how did the sequence come to encode a smooth folding funnel in the first place?" Evolutionary searches through sequence space face the same combinatorial explosion as conformational searches — with the additional constraint that only functional sequences are selectable.
Chaperones add a further layer of irreducible complexity: they are themselves folded proteins that must have been folded correctly in the first place. The system that assists folding requires the very folding it assists — a chicken-and-egg dependency that makes gradual evolutionary emergence of the system impossible to reconstruct stepwise.